

Publicaciones SERIDA
Nuevos marcadores de calidad de madera en Pinus pinaster: Estrigolactonas y ramificación.
ÁLVARO CALDERÓN. Universidad de Oviedo, Departamento Biología de Organismos y Sistemas, Área de Fisiología Vegetal.
ISABEL FEITO. Área de Cultivos Hortofrutícolas y Forestales, Programa Forestal. ifeito@serida.org
CAROLINA DE LA TORRE. Área de Cultivos Hortofrutícolas y Forestales, Programa de Investigación Forestal.
JESÚS PASCUAL. Universidad de Oviedo, Departamento Biología de Organismos y Sistemas, Área de Fisiología Vegetal.
FRANCISCO JAVIER COLINA. Universidad de Oviedo, Departamento Biología de Organismos y Sistemas, Área de Fisiología Vegetal.
LUIS VALLEDOR. Universidad de Oviedo, Departamento Biología de Organismos y Sistemas, Área de Fisiología Vegetal.
JUAN MAJADA. Centro Tecnológico Forestal y de la Madera de Asturias, Programa de Investigación Forestal
ANA RODRÍGUEZ. Universidad de Oviedo, Departamento Biología de Organismos y Sistemas, Área de Fisiología Vegetal.
MARÍA JESÚS CAÑAL. Universidad de Oviedo, Departamento Biología de Organismos y Sistemas, Área de Fisiología Vegetal.
MÓNICA MEIJÓN. Área de Cultivos Hortofrutícolas y Forestales, Programa Forestal. mmeijon@serida.org
Introducción
El consumo de madera y de subproductos de origen forestal ha experimentado un gran incremento durante los últimos años (basado en FAO 2008c), haciendo que los productos derivados de la explotación forestal tengan cada vez un mayor valor añadido. Estos materiales renovables se encuentran, tanto a nivel europeo como nacional, en el foco de diferentes planes de acción que se fundamentan en su potencial sostenibilidad en el marco de un clima cambiante y su posición de activo estratégico por las posibilidades que ofrece en la mejora de la autosuficiencia y disponibilidad estable de materiales más ecológicos. Por todo ello, se financia la investigación y mejora de la explotación en torno a este recurso, con distintos incentivos como por ejemplo el programa Horizonte 2020, “Raw Materials”. Las coníferas, y en concreto el genero Pinus, adquiere en este marco un claro protagonismo a nivel nacional, al constituir hoy en día el principal recurso de abastecimiento de la industria de aserrado en la Península Ibérica. Una de las singularidades que presentan la mayoría de las coníferas es su particular patrón de crecimiento puesto que, al contrario de otras especies forestales, son capaces de dar lugar a la formación y elongación de más de una unidad de tallo en un mismo período anual (varios ciclos de crecimiento), siempre y cuando las condiciones ambientales sean favorables. Este comportamiento denominado policiclismo se produce en especies del género Pinus, como el Pinus pinaster, de gran interés no solo para la industria de primera y segunda transformación, sino también a nivel ecológico. Este interés se debe fundamentalmente a su amplia distribución natural en la Península Ibérica, a su capacidad de crecimiento en suelos pobres y bajo periodos de sequía prolongada (Alía y Martin. 2009) y a la gran diversidad genética de la especie, que da lugar a diferencias de comportamiento en función de la procedencia de la población. En España, es el género Pinus el que ocupa de forma natural la mayor superficie, y en Asturias llega a valores próximos al 10 % del total de la superficie forestal.
La industria del aserrado es la actividad económica más importante en la transformación del P. pinaster no sólo por su elevado nivel de consumo de materia prima (67 % del total), sino también por su incidencia como fuente de abastecimiento de productos y subproductos destinados a otros subsectores. Algunos de estos sectores utilizan preferentemente madera de alto valor comercial, sin nudos y homogénea. Sin embargo, este tipo de madera no es fácil de obtener debido a la ausencia de gestión y la alta proclividad de esta especie a un modelo de crecimiento policíclico, lo que hace que los individuos muestren una gran ramificación y, por tanto, un gran número de nudos. Este carácter presenta una gran heredabilidad (Zas y Fernández-López, 2005); sin embargo se desconocen por completo las bases moleculares que inducen su manifestación.
La ramificación del tallo está controlada por la formación y subsecuente crecimiento de yemas axilares, que son estructuras indeterminadas cuyo crecimiento se activa por señales endógenas o ambientales. Más de 70 años después de que se propusiera que la fitohormona auxina, sintetizada en la zona apical de las plantas, actuaba como un inhibidor de la ramificación (Thimann y Skoog 1933), y tras la teoría propuesta del segundo mensajero (Brewer et al., 2009), estudios recientes han descubierto que un grupo específico de lactonas terpenoides denominadas estrigolactonas pueden inhibir directa o indirectamente el crecimiento de la yema.
¿Qué son las estrigolactonas?
Las estrigolactonas, son un nuevo grupo de hormonas vegetales, cuyo papel era desconocido hasta hace poco. Desde mediados del siglo XX se sabía que dicha familia de compuestos se producía en las raíces de las plantas y se exudaba en la rizosfera para «atraer» hongos y facilitar el desarrollo de simbiosis endomicorricica (Cook et al., 1972). Sin embargo, han tenido que transcurrir 50 años para que se localice la actividad endógena de estas hormonas dentro de las plantas (Beveridge y Hyozuka, 2010), relacionando su actividad con la inhibición de la ramificación. Este tipo de hormonas poseen ciertas características comunes al resto de las fitohormonas: su acción es muy específica, son activas en concentraciones muy bajas y pueden recorrer largas distancias en la planta. Trabajos realizados en la última década han hecho posible alcanzar un mayor conocimiento sobre la función de estas fitohormonas en la ramificación. Además, recientemente se han identificado nuevos genes implicados en la ramificación de los brotes y la biosíntesis de estrigolactonas en diversas especies modelo, como arabidopsis, guisante, arroz y petunia (Brewer et al., 2009). No obstante, aún son escasos los trabajos en especies forestales relacionados con este campo.
A través de recientes estudios (Booker et al., 2005; Brewer et al., 2009; Domagalska and Leyser, 2011), se ha podido comprobar que la ruta de biosíntesis y señalización de estrigolactonas está ampliamente conservada entre distintas especies vegetales, especialmente la superfamilia MAX (MORE AXILLARY GROWTH). Según el modelo de inhibición de la ramificación vía estrigolactonas (Figura 1), la biosíntesis del precursor de estrigolactonas es mediada por dos CAROTENOID CLEAVAGE DIOXYGENASES (CDD) codificadas por MAX3 (CCD7) y MAX4 (CCD8). Tras esto, el precursor de estrigolactonas, la carlactona, es transformada bioquímicamente a través del citocromo P450 (MAX1) en una estrigolactona bioactiva móvil. A continuación, en la yema lateral, y mediante el receptor de la señalización de estrigolactonas (MAX2), se inhibe la activación del crecimiento de la yema lateral. Las auxinas juegan también un papel esencial en este entramado, ya que interfieren en la biosíntesis de estrigolactonas mediante la vía de transducción de señales regulada por AUXIN RESISTAN1 (AXR1). Por otro lado, además, AXR1 inhibe la biosíntesis de citoquininas, suprimiendo, por consiguiente, la formación de nuevas yemas.
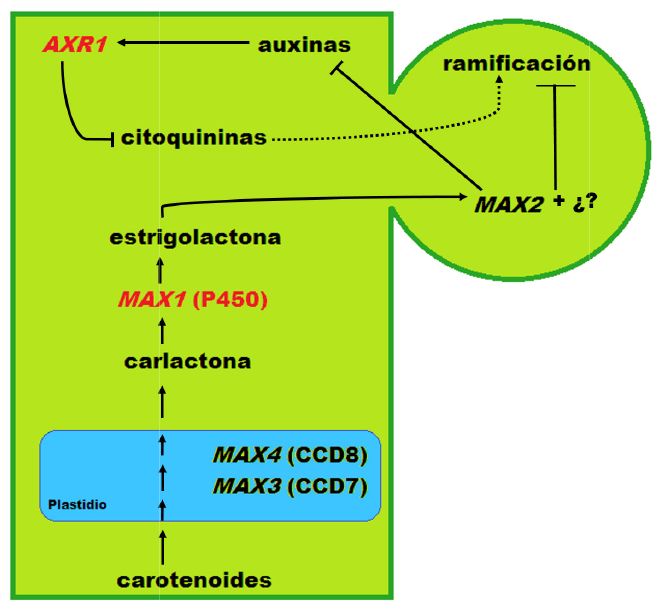
Figura 1. Modelo simplificado de la inhibición de la ramificación de los brotes laterales vía estrigolactonas. MAX: MORE AXILLARY GROWTH; CCD: CAROTENOID CLEAVAGE DIOXYGENASES; AXR1: AUXIN RESISTANT 1; P450: citocromo P450; ¿? = otros factores desconocidos. En rojo los genes estudiados en este trabajo.
Primeras pistas del papel de AXR1 y MAX1 en P. pinaster
Son muy pocos, los trabajos existentes hasta la fecha dirigidos al estudio del posible papel de las estrigolactonas en la regulación de la ramificación y arquitectura de las especies forestales. Es por eso que inicialmente se decidió realizar un análisis filogénetico de los principales genes implicados en la biosíntesis de estrigolactonas y su señalización en diez especies secuenciadas, incluyendo entre ellas varias especies forestales, para determinar el grado de conservación de esta ruta de señalización y establecer los genes diana más interesantes en nuestro trabajo, antes de analizar el papel específico de estas hormonas en el control de la ramificación de P. pinaster.
Mediante este estudio filogenético (Figura 2) se pudo observar que AXR1 es el gen más variable entre las especies estudiadas, mientras que la superfamilia MAX se encuentra altamente conservada incluso entre especies tan distantes como, P. taeda y Arabidopsis thaliana. Con esta información, y con la idea de avanzar en el conocimiento acerca de los factores endógenos implicados en la regulación del policiclismo y ramificación en coníferas, se decidió estudiar el posible papel de los siguientes genes en la inhibición de la ramificación de P. Pinaster: AXR1, por ser el gen que mostró mayor variabilidad entre especies, y dentro de la familia MAX, altamente conservada, MAX1, por ser el gen de mayor relevancia en la ruta de biosíntesis de estrigolactonas, ya que su función es clave en la movilización y activación de las estrigolactonas. Partiendo de la información disponible sobre el genoma de P. pinaster (Canales et al., 2014) y P. taeda (Neale et al., 2014), y mediante el diseño de cebadores degenerados, fue posible obtener la secuencia homóloga de ambos genes en P. pinaster.
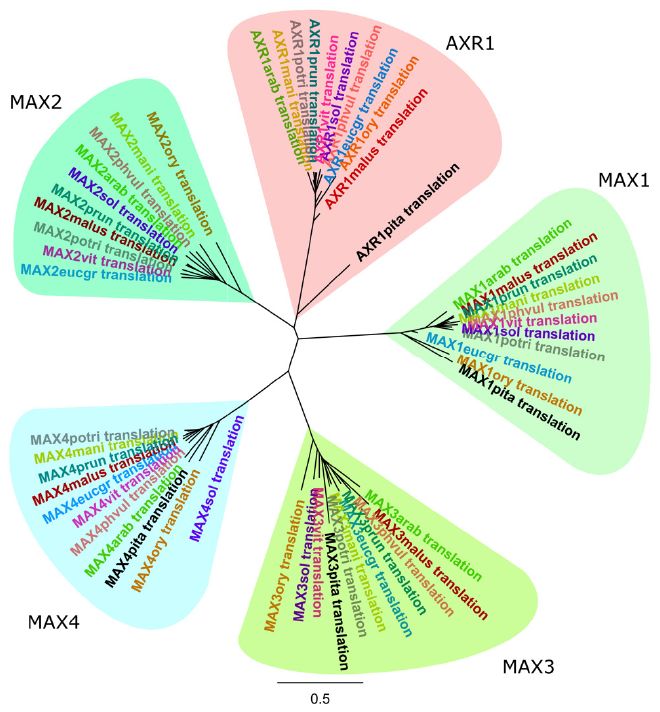
Figura 2. Dendrograma de los principales genes relacionados con la biosíntesis y señalización de estigolactonas mostrando las relaciones de similitud existentes entre las distintas especies secuenciadas seleccionadas. Abreviaturas de las especies utilizadas: arab, Arabidopsis thaliana; mani, Manihot esculenta; potri, Populus trichocarpa; prun, Prunus persica; vit, Vitis vinífera; sol, Solanum tuberosum; phvul, Phaseolus vulgaris; eucgr, Eucalyptus grandis; ory, Oryza sativa; malus, Malus domestica; pita, Pinus taeda. Ejemplo nomenclatura en el dendograma: MAX4potri translation, proteína traducida de MAX4 en la especie Populus trichocarpa.
Con el objeto de establecer el papel fisiológico de estas fitohormonas en la inhibición del crecimiento policíclico, se llevaron a cabo varios ensayos empleando como material vegetal plántulas de P. pinaster de 2 años de edad, bifurcadas (con sólo una rama lateral; Figura 3). En el primero de los ensayos se aplicaron los siguientes tratamientos en diferentes combinaciones: estrigolactona sintética (GR-24), un inhibidor del transporte de auxinas (ácido naftil-p-talámico, NPA), citoquininas (benziladenina, BA) y poda de rama dominante. Tras este primer estudio, se comprobó que los resultados más prometedores en el control de la ramificación se alcanzaban en aquellos tratamientos en los que se había aplicado GR-24 y/o NPA, por lo que se decidió realizar un segundo ensayo más restrictivo, en el que se aplicaron los dos compuestos sobre la rama lateral y/o la rama dominante en diferentes combinaciones (Figura 3). Los datos fenológicos muestran diferencias significativas entre los distintos tratamientos, observándose que las plántulas tratadas con GR-24 sobre la rama lateral (tto.3) son las que presentan mayor crecimiento en altura, además de una clara disminución del crecimiento en la rama lateral. Por otro lado, la aplicación del inhibidor del transporte de auxinas (NPA), independientemente de sobre qué rama se realice (dominante o lateral), provoca una disminución en el crecimiento de la rama lateral (ttos. 4, 5 y 7; figura 3). Tan sólo en el caso de la aplicación de NPA sobre la rama dominante, cuando se aplica además GR-24 sobre la misma rama (tto. 6), conlleva que se pierda el efecto de inhibicioìn del crecimiento en la rama lateral provocado por este quiìmico; sin embargo, en este caso no se aprecia un incremento significativo respecto al control en la altura de la rama dominante.
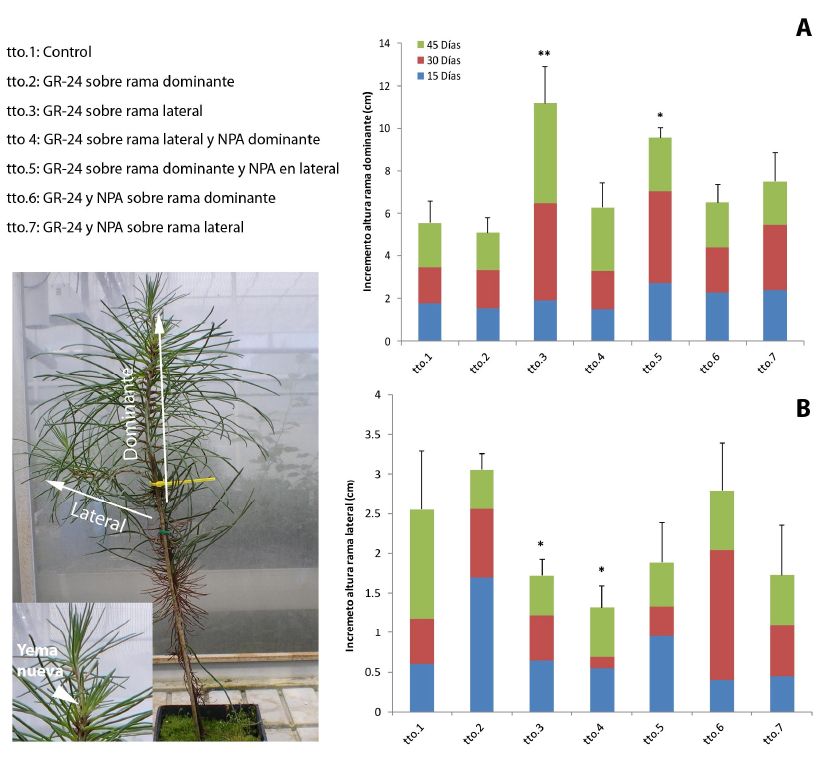
Figura 3. Columna izquierda, tratamientos aplicados y ejemplo de plántula de Pinus pinaster bifurcada sobre la que se marcan las ramas dominante y lateral. Columna derecha, datos de crecimiento de las plántulas bifurcadas. (A) Incremento en altura de la rama dominante a los 15, 30 y 45 días de la aplicación de los tratamientos. (B) Incremento en altura de la rama lateral a los 15, 30 y 45 días de la aplicación de los tratamientos. T-Student comparando cada uno de los tratamientos frente al control (tto.1). Los asteriscos indican diferencias significativas (** p ≤ 0,01; * p ≤ 0,05; n = 10). GR-24: Estrigolactona sintética; NPA = ácido naftil-p-talámico, inhibidor del transporte de auxinas; tto. Tratamiento.
La obtención de las secuencias de los genes homólogos a AXR1 y MAX1 en P. pinaster (PpAXR1 y PpMAX1), permitió realizar un análisis de expresión de ambos genes en los diferentes tratamientos, tanto en las yemas laterales como dominantes, lo que en último termino nos permitirá determinar la funcionalidad de la ruta de señalización de las estrigolactonas en el control de la inhibición de la ramificación en P. pinaster. Así, se observó que las yemas dominantes tratadas con el NPA muestran una inhibición de la expresión tanto de PpAXR1 como de PpMAX1, mientras que las yemas laterales tratadas con GR-24 presentan los mayores niveles de expresión para ambos genes. A través de estos resultados se logra establecer, por primera vez, una relación directa entre la actividad de ambos genes y la inhibición de la ramificación, puesto que estas plántulas, que experimentan un incremento en la expresión de estos genes, son las que experimentaron también el mayor incremento en altura de la rama dominante y el menor en la rama lateral (tto. 3). Además, se validaron estos resultados mediante hibridación in situ fluorescente, visualizándose la expresión de ambos genes en el tejido y revelando así la localización exacta de su actividad dentro de las plantas. De este modo, se apreció que, aunque la expresión de ambos genes se localiza principalmente en la yema lateral de los tratamientos con mayor inhibición de la ramificación (tto.3), PpAXR1 presenta una expresión apical, mientras que la expresión de PpMAX1 se localiza preferentemente en la parte basal de la yema (Figura 4).
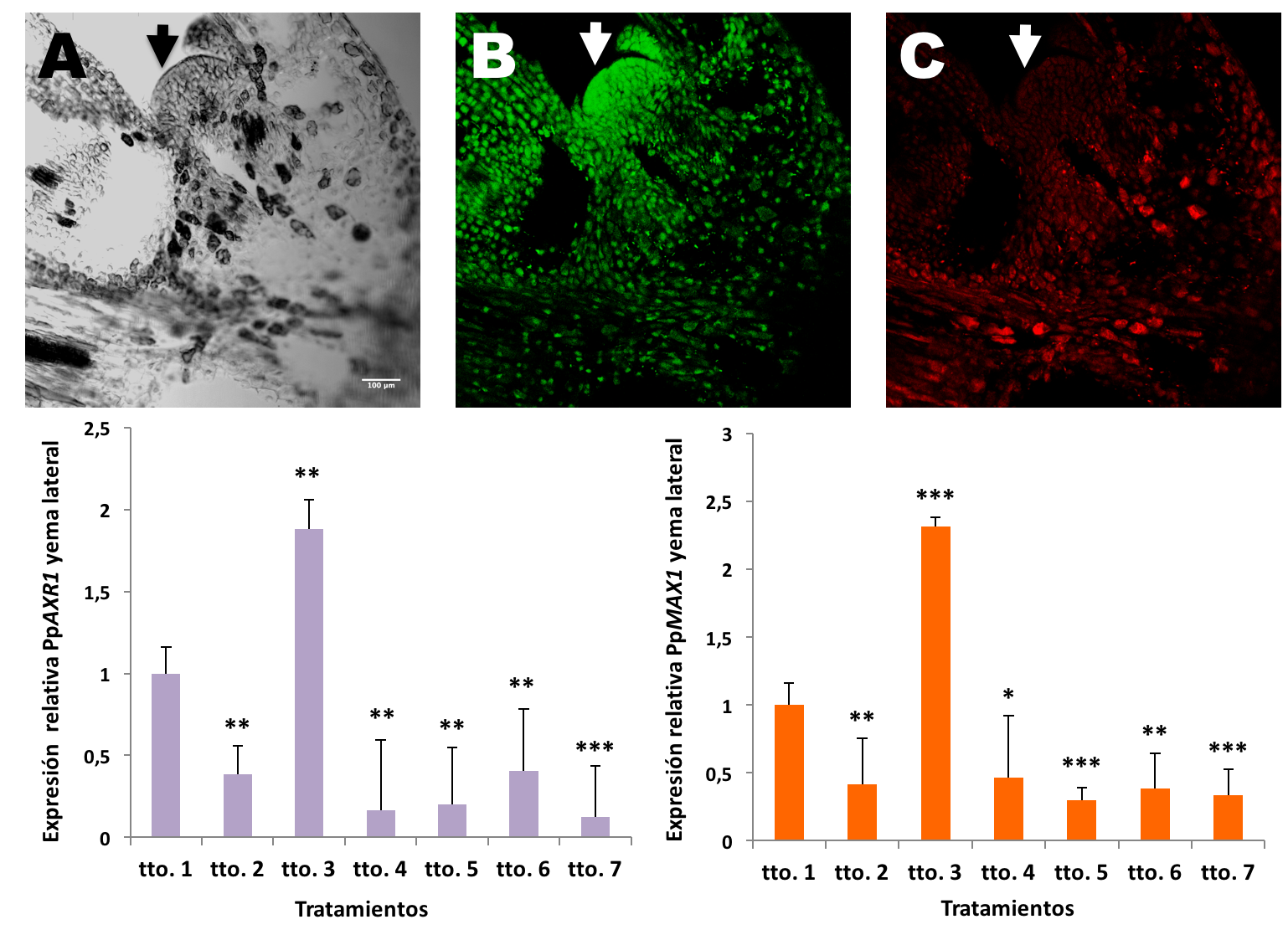
Figura 4. Parte superior, FISH en yemas laterales de Pinus pinaster del tto. 3; (A) Microscopía de contraste por interferencia diferencial (DIC) con un objetivo de inmersión 20x en yemas laterales; (B) Expresión de PpAXR1; (C) Expresión de PpMAX1. La flecha blanca indica la posición del meristemo. Abreviaturas: tto. 3 = adición de GR-24 sobre la rama lateral. Parte inferior, expresión de PpAXR1 y PpMAX1 en yemas laterales de los distintos tratamientos 30 días después de su aplicación. Datos de expresion relativa normalizados frente al control (tto.1). T-Student comparando cada unos de los tratamientos frente al control (tto.1). Los asteriscos indican diferencias significativas (*** p ≤ 0,001; ** p ≤ 0,01; * p ≤ 0,05; n = 3). tto. 1: control; tto. 2: GR-24 sobre la rama dominante; tto. 3: GR-24 sobre la rama lateral; tto. 4: GR-24 en rama lateral y NPA en rama dominante; tto. 5: GR-24 en rama dominante y NPA en rama lateral; tto. 6: GR-24 y NPA en rama dominante; tto. 7: GR-24 y NPA en rama lateral.
En base a estos resultados, se puede afirmar que la función de las estrigolactonas en la inhibición de la ramificación en P. pinaster se ejerce fundamentalmente a nivel de las yemas laterales, lo cual repercute en un mayor incremento en longitud de la rama dominante. Por otro lado, según los resultados del tratamiento con NPA, se puede deducir que las auxinas juegan también un papel decisivo en la regulación de la actividad de las estrigolactonas en la inhibición de la ramificación, ya que el tratamiento con estrigolactonas (GR-24) sobre yema lateral en plantas previamente tratadas con NPA (inhibidor del transporte de auxinas) no muestra diferencias significativas respecto al control, ni a nivel fenológico, ni a nivel de expresión de PpAXR1 o PpMAX1.
Relevancia de la ruta de señalización de las estrigolactonas en el diseño de los nuevos programas de mejora forestal
En la parcela experimental de “La Mata“, en Grado, el SERIDA dispone de una colección clonal que presenta una estructura genética de procedencias-progenie de P. pinaster, la cual cubre toda la variación natural de la especie. En el seguimiento de la colección en campo se ha observado que las procedencias muestran diferente grado de policiclismo y, en consecuencia, distintos hábitos de ramificación en relación a su origen: Mediterráneo o Atlántico.
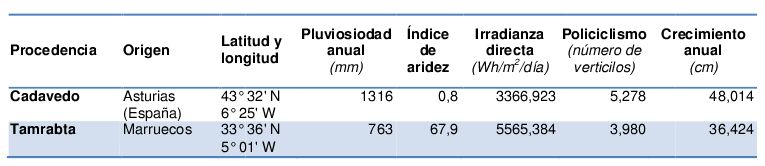
Partiendo de este material, y con la información adquirida en los ensayos descritos previamente, se decidió llevar a cabo la cuantificación de la expresión de PpAXR1 y PpMAX1 en dos procedencias de P. pinaster que presentan un crecimiento policíclico altamente contrastante: Tamrabta (Marruecos) y Cadavedo (Asturias-España) (Tabla 1). De este modo se observó que ambos genes presentan altos niveles de expresión en la procedencia Tamrabta, coincidiendo esto con el fenotipo de esta población, el cual muestra baja incidencia en crecimiento policíciclo; mientras que la menor expresión, para ambos genes, se detecta en la procedencia Cadavedo (Asturias), la población que muestra el mayor grado de ramificación y policiclismo.
Tabla 1. Origen y detalles climáticos y de crecimiento de las procedencias incluidas en el estudio. Los datos climáticos fueron tomados de Sánchez-Gómez et al. (2010), los de irradiación de “European Climate Assessment & Dataset”, y los datos de crecimiento y policiclismo fueron adquiridos por el SERIDA en septiembre de 2013 en la colección clonal de Pinus pinaster establecida en la finca experimental “La Mata” (Grado).
En este trabajo se demuestra, por primera vez, la implicación de las estrigolactonas en la regulación de la ramificación de P. pinaster. Los ensayos establecidos, así como los resultados de expresión de los genes estudiados en los árboles de distintas procedencias, aportan la primera pista acerca de la relevancia de las estrigolactonas en la inhibición del policiclismo en esta especie. La implementación de estos resultados en los futuros programas de mejora de P. pinaster permitirá mejorar la calidad y la productividad de esta especie. El uso de las estrigolactonas como marcadores tempranos de desarrollo facilitará la identificación, y posterior multiplicación clonal, de aquellos individuos que posean características de interés: árboles que presenten una mayor ramificación para la producción de biomasa con fines energéticos, o árboles que presenten menor ramificación para la producción de madera sin nudos y de alto valor comercial.
Agradecimientos
Este trabajo se ha podido realizar gracias a la colaboración entre el Área de Fisiología Vegetal de la Universidad de Oviedo y el Programa Forestal del SERIDA (Servicio Regional de Investigación y Desarrollo Agroalimentario del Principado de Asturias). Este trabajo forma parte de los Proyectos Fin de Grado en Biología y Fin de Máster de Álvaro Calderón González, dentro del Máster en Biotecnología Aplicada a la Conservación y Gestión Sostenible de Recursos Vegetales, y se engloba dentro del Proyecto RTA 2013-00048-C03-02, Subprograma de Proyectos de Investigación Fundamental Orientada a los Recursos y Tecnologías Agrarias obtenido por el Programa de Investigación Forestal del SERIDA.
Referencias Bibliográficas
ALÍA, R., MARTÍN, S. (2009). Guía técnica para la conservación genética y utilización del pino negral (Pinus pinaster) en España. Foresta. Madrid. España.
BEVERIDGE, C.A., HYOZUKA, J. (2010). New genes in the strigolactones-related shoot branching pathway. Curr Opin Plant Biol. 13, 34-39.
BOOKER, J., SIEBERER, T., WRIGHT, W., WILHAMSON, L., WILLETT, B., ET AL. (2005). MAX1 encodes a cytochrome P450 family member that acts downstream of MAX3/4 to produce a carotenoid-derived branch-inhibiting hormone. Dev Cell. 8, 443-449.
BREWER, P.B., DUN, E.A., FERGUSON, B.J., RAMEAU, C., BEVERIDGE, C.A. (2009). Strigolactone acts downstream of auxin to regulate bud outgrowth in pea and Arabidopsis. Plant Physio., 150, 482-493.
CANALES, J., BAUTISTA, ROCIO., LABEL, P., GÓMEZ-MALDONADO, J., LESUR, I., FERNÁNDEZ-POZO, N., RUEDA-LÓPEZ, M., GUERRERO-FERNÁNDEZ, D., CASTRO-RODRIGUEZ, V., BENZEKRI, H., CAÑAS, R.A., GUEVARA, M.A., RODRIGUES, A., SEOANE, P., TEYSSIER, C., MOREL, A., EHRENMANN, F., LE PROVOST, G., LALANNE, C., NOIROT, C., KLOPP, C., REYMOND, I., GARCÍA-GUTIERREZ, A., TRONTIN, J.F., LELU-WALTER, M.A., MIGUEL, C., CERVERA, M.T., CANTÓN, F.R., PLOMION, C., HARVENGT, L., AVILA, C., CLAROS, M.G., CÁNOVAS, F.M. (2014). De novo assembly of maritime pine transcriptome: implications for forest breeding and biotechnology. Plant Biotechnology Journal.
COOK, C.E., WHICHARD, L.P., WALL, M.E., EGLEY, G.H., COGGON, P., LUHAN, P.A., MCPHAIL, A.T. (1972). Germination stimulants. II. The structure of strigol: a potent seed germination stimulant for witchweed (Striga lutea Lour.). J Am Chem Soc., 94, 6198-6199.
DOMAGALSKA, M.A., LEYSER, O. (2011). Signal integration in the control of shoot branching. Nat Rev Mol Cell Biol. 12, 211-221.
GOMEZ-ROLDAN, V., FERMAS, S., BREWER, P.B., PUECH-PAGES, V., DUN, E.A., PILLOT, J.P., LETISSE, F., MATUSOVA, R., DANOUN, S., PORTAIS, J.C., BOUWMEESTER, H., BECARD, G., BEVERIDGE, C.A., RAMEAU, C., ROCHANGE, S.F. (2008). Strigolactone inhibition of shoot branching. Nature, 455, 189-194.
NEALE, D.B., WEGRYN, J.L., STEVENS, K.A., ZIMIN, A.V., PUIU, D., CREPEAU, M.W., CARDERO, C., KORIABINE, M., HOLTZ-MORRIS, A.E., LIECHTY, J.D., MARTÍNEZ-GARCÍA, P.J., VASQUEZ-GROSS, H.A., LIN, B.Y., ZIEVE, J.J., DOUGHERTY, W.M., FUENTES-SORIANO, S., WU, L.S., GILBERT, D., MARCAIS, G., ROBERTS, M., HOLT, C., YANDELL, M., DAVIS, J.M., SMITH, K.E., DEAN, J.FD., LORENZ, W.W., WHETTEN, R.W., SEDEROFF, R., WHEELER, N., MCGUIRE, P.E., MAIN, D., LOOPSTRA, C.A, MOCKAITIS, K., DEJONG, P.J., YORKE, J.A., SALZBERG, S.L., LANGLEY, C.H. (2014). Decoding the massive genome of loblolly pine using haploid DNA and novel assembly strategies. Genome Biology.
SÁNCHEZ-GÓMEZ, D., MAJADA, J., ALÍ, R., FEITO, I., ARANDA, I. (2010). Intraspecific variation in growth and allocation patterns in seedlings of Pinus pinaster Ait. submitted to contrasting watering regimes: can water availability explain regional variation?. Ann. For. Sci. 67, 505.
THIMANN, K.V., SKOOG F. (1933). Studies on the growth hormone of plants: III. The inhibiting action of the growth substance on bud development. Proc. Nati Acad. Sci. U.S.A., 19, 714-716.
ZAS, R., FERNÁNDEZ-LÓPEZ, J. (2005). Juvenile genetic parameters and genotypic stability of pinus pinaster Ait. Open-pollinated families under different water and nutrient regimes. Forest Science,51(2), 165-174(10).
| Ficha Bibliográfica | |
|---|---|
| Título | Nuevos marcadores de calidad de madera en Pinus pinaster: Estrigolactonas y ramificación. |
| Autor/es | ÁLVARO CALDERÓN. Universidad de Oviedo, Departamento Biología de Organismos y Sistemas, Área de Fisiología Vegetal. ISABEL FEITO. Área de Cultivos Hortofrutícolas y Forestales, Programa Forestal. ifeito@serida.org CAROLINA DE LA TORRE. Área de Cultivos Hortofrutícolas y Forestales, Programa de Investigación Forestal. JESÚS PASCUAL. Universidad de Oviedo, Departamento Biología de Organismos y Sistemas, Área de Fisiología Vegetal. FRANCISCO JAVIER COLINA. Universidad de Oviedo, Departamento Biología de Organismos y Sistemas, Área de Fisiología Vegetal. LUIS VALLEDOR. Universidad de Oviedo, Departamento Biología de Organismos y Sistemas, Área de Fisiología Vegetal. JUAN MAJADA. Centro Tecnológico Forestal y de la Madera de Asturias, Programa de Investigación Forestal ANA RODRÍGUEZ. Universidad de Oviedo, Departamento Biología de Organismos y Sistemas, Área de Fisiología Vegetal. MARÍA JESÚS CAÑAL. Universidad de Oviedo, Departamento Biología de Organismos y Sistemas, Área de Fisiología Vegetal. MÓNICA MEIJÓN. Área de Cultivos Hortofrutícolas y Forestales, Programa Forestal. mmeijon@serida.org |
| Año Publicación | 2016 |
| Área | |
| Revista/Serie | Tecnología Agroalimentaria. Boletín informativo del SERIDA |
| Referencia | nº 17, pp. 21 - 27 |
| Formato | |
| Depósito Legal | As.-2.617/1995 |
| ISSN | 1135-6030 |
| ISBN | |
| Ver/Descargar | http://www.serida.org/publicacionesdetalle.php?id=6362&anyo= |